Molecular Mind in Cellular Motors 6: The Very Talented Dynein Motor
- Jon Lieff

- May 28
- 5 min read

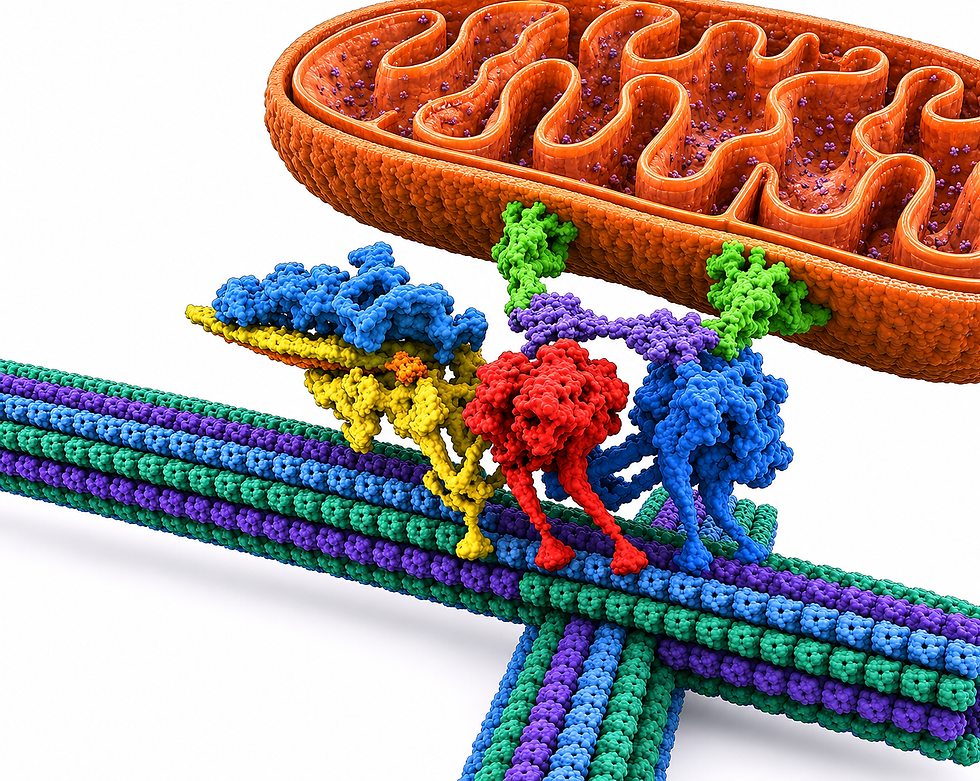
Dynein is a vastly more complex motor than kinesin. It walks along microtubules carrying vesicles, RNA granules, mitochondria, signaling complexes, and even large full chromosomes. It moves the entire cell nucleus during immune cell movement and manages the extraordinarily complex choreography of chromosome separation during cell division. It is a motor with awareness of what each situation requires.
Dynein is enormous. Including its essential additional dynactin complex, it has 21,000 amino acids, compared to kinesin’s 3,000. This is larger than a ribosome, one of the largest molecular clusters. Dynein has twelve subunits made of heavy, intermediate, light intermediate, and light chains, as well as the associated dynactin complex, which has an additional twenty subunits.
.
Dynein operates everywhere in the cell using essentially one universal molecular design, while kinesin has diversified into 45 types, each specialized for particular cargo and cell type. Dynein, like kinesin, is assembled through weak non-covalent bonds, giving the entire structure the flexibility it needs to walk, bend, and adapt.
Unlike kinesin's single, uniform step, dynein walks in many modes. It can walk exactly like a human, alternating left and right feet in equal steps. It can move like an inchworm, with one foot consistently leading while the other follows. It can take repeated steps with one foot before the other moves at all. It can step sideways off one microtubule track and onto a parallel or perpendicular one. This multi-gait capability allows dynein to navigate complex three-dimensional scaffold environments, carry diverse cargo of wildly different sizes and shapes, circumvent large obstacles by rotating around or stepping off the track entirely, and operate across a far wider range of cellular contexts than kinesin alone could manage.

Its step sizes vary enormously—the largest step is at least eight times the size of the smallest, with steps along the tubulin-based track ranging from half a tubulin unit to the length of four tubulin units. This variability is made possible by dynein's long flexible stalk, which can swivel in multiple directions to reach different attachment sites on the microtubule, and by the large arc of its linker arm, whose trajectory on each swing determines both the size and the direction of the step. The linker arm in dynein is substantially larger than kinesin's equivalent linker, and its wide arc of movement is what gives dynein its extraordinary range of step sizes and walking styles.
Dynein's complex motor is built around a ring of six enzymes in the heavy chain, each of which is designed to use ATP. This ring is a molecular engine that changes shape as it cycles through energy reactions. The first enzyme in the ring drives most of the stepping, while others serve as sensors, anchors, regulators, and emergency backups. The ring changes shape at each stage of the chemical reaction that breaks ATP. Those shape changes are transmitted through a large flexible linker arm that drives the stepping motion. The ring's shape shifts to tell the motor how heavy its load is, how far to step, and whether to pause.

ATP binds to the ring enzyme and that chemical reaction breaks ATP into two parts releasing a large amount of energy. This energy is transduced into mechanical movement. Each ATP reaction shifts the geometry of the ring. Dynein’s adaptive behavior is mediated by a cascade of non-covalent conformational changes that ripple through the protein. Every step dynein takes involves waves of changing non-covalent interactions spreading from its giant ring of ATPase enzymes to the linker domain, then to the stalk, and finally to the microtubule track binding domain.
Dynein has multiple operating modes that adapt continuously to the actual conditions of each transport job, based on feedback. Dynein’s internal tension-sensing architecture is what allows a single motor type to handle everything from rapid organelle delivery to the slow, forceful repositioning of the entire cell nucleus during immune cell movement. Dynein’s internal communication system is remarkably sophisticated. Only one of the six enzymes reacts with ATP. The other enzymes in the ring serve as mechanical sensors, anchored in place to detect strain and tension as the motor walks. When the shape of the ring shifts, these sensors register the change and alter the timing of ATP reactions, which adjusts step length, step timing, and force output in real time––e.g., when one foot is carrying an unusually heavy load the sensors can hold it steady while the other foot inches forward.
Dynein is unique among molecular motors in that it cannot move at all without its companion, the massive dynactin complex. Dynactin is built around a central filament made of eight identical connected subunits, with a pointed end cluster of three molecules and a side arm of nine molecules. Dynactin assembles in a precise sequence and once assembled, it binds to dynein's tail at two contact points and recruits cargo adaptor molecules, forming the complete active dynein complex.
The dynactin complex connects dynein to its cargo. It also physically straightens the two dynein heavy chains from their inactive, folded configuration into an open, parallel alignment that allows the feet to attach to the microtubule and walk. Without dynactin, dynein cannot activate. Sometimes, dynactin incorporates a second complete dynein motor alongside the first, dramatically increasing the speed and force available for carrying large or heavy loads. Dynactin is not a passive scaffold—it is an active regulator, communicating continuously with dynein's motor domains throughout every step.

Dynein is not simply a motor that burns ATP, but a molecular creature that continuously senses, adapts, negotiates forces, and changes behavior depending on context. It can switch directions, coordinate with kinesin, avoid obstacles on crowded microtubules, regulate force production, and synchronize with multiple partner proteins. Dynein behavior is due to its weak non-covalent interactions, the changing of surrounding water hydration shells, and actions from disordered protein regions. Dynein’s huge complex structure constantly fluctuates, with its motor never existing in one fixed shape. Instead, it tests multiple different conformations, with information flowing through shifting patterns of weak bonds. The changing hydration shells act like computational fields that shape timing and coordination by stabilizing some protein states and destabilizing others. Every time water networks reorganize, they in turn influence dynein’s next shape, and therefore its actions.
Dynein and kinesin do not simply carry packages from one place to another—they are nodes in a cell-wide communications and logistics network of extraordinary complexity involving a large society of individual molecular creatures. Every aspect of this operation involves continuous signaling among these individual creatures. Chemical tags are added to or removed from motors, adaptors, tracks, and cargo. Mechanical forces are transmitted between motors sharing the same load. Shape changes propagate through entire molecular complexes. Electrical charge distributions on the microtubule surface act as traffic signs, routing motors onto specific tracks or away from them.
How can we not consider these molecular creatures intelligent?